Introducción: Sobre los organismos-modelo en estudio con plantas.
En la novedosa área de estudio sobre la cognición de las plantas, destinada a evaluar la posibilidad sobre si las plantas pueden ser agentes sensibles (e.g Bohm et al. 2016; Calvo et al. 2017; Dudley y File 2007) y cognitivas (e.g. Gagliano et al. 2016; Gagliano 2017; Segundo-Ortin y Calvo 2021), uno de los problemas que parecen suscitarse es relativo a la pregunta sobre cómo los investigadores, es decir, quienes observan y estudian a estos organismos, son capaces de presenciar las características y comportamientos que se identifican con la existencia de estos fenómenos biológicos. Lo habitual es que estos organismos no muestren comportamientos propios de organismos sensibles y cognoscentes como los animales, o más bien, parece que no los muestran debido a sus movimientos excesivamente lentos que pueden resultar imperceptibles para el ser humano (Calvo et al. 2017). Por ello, resulta relevante analizar con qué sujeto de estudio o con qué organismo-modelo se está evaluando la presencia del fenómeno sensitivo y cognitivo en plantas.
A este efecto, una forma particular de hacer investigación en ciencias es con los llamados organismos- modelo. Estos son definidos como “especies no humanas que se estudian ampliamente para comprender
una variedad de fenómenos biológicos” (Ankeny y Leonelli 2020, 2). Generalmente en biología, botánica o simplemente en estudios del reino vegetal, el organismo-modelo más utilizado es la hierba Arabidopsis Thaliana. Esta es utilizada debido a su baja estatura, lo que la hace fácilmente manipulable, y también por su rápido crecimiento o generación, lo cual facilita el estudio rápido de los genotipos (Meinke et al. 1998). Gracias a ella se han podido cartografiar diversas mutaciones fenotípicas y bioquímicas que permiten analizar a los organismos vegetales (Schmid et al. 2005). Otro organismo- modelo que resulta interesante en los estudios de genoma y herencia genética es la famosa Pisum Sativum, la planta de arvejas utilizada por Mendel (Weeden 2016). Dicha planta no solo ha participado como modelo en cuestiones relativas a la genética, sino que también ha participado en experimentos relativos al aprendizaje en plantas. En Gagliano et al. (2016) se evidencia como este modelo es capaz de aprender a través de asociación de estímulos, permitiendo postular que las plantas son organismos capaces de manipular información por ciertos periodos de tiempo en beneficio de su sobrevivencia.
Con los organismos-modelo se tiene la pretensión de proporcionar cierta comprensión sobre otros organismos de la misma especie (Ankeny y Leonelli 2020,2), por lo que, a partir de estudios como los de Gagliano et al. (2016) podríamos concluir que, en algún grado, todos los demás organismos vegetales cuentan con algún tipo de mecanismo que les permite recopilar información del medio y utilizarla para aprender de él y aumentar sus probabilidades de sobrevivencia.
En estudios sobre la sensibilidad y la cognición en plantas no parece existir un modelo acordado para estudiar estos fenómenos. Podría resultar sugerente plantear que algún proceso sensible o cognitivo es inexistente en plantas porque la mayoría de éstas parecen no moverse. Autores como Burdon-Sanderson (Shepherd 2012) plantean que solo las plantas que inician movimientos visibles para el ser humano, tales como Mimosa Púdica o Dionaea Muscipula, son plantas sensibles y, por lo tanto, ser potencialmente agentes cognitivas. Sin embargo, sabemos que todas las plantas inician movimientos, solo que no todas son observables por el ser humano (Calvo et al. 2017): esto es fácilmente contrastable mediante técnicas de aceleración de fotogramas llamadas timelapse.
Mimosa Púdica es una planta de movimientos rápidos que son directamente observables por el ser humano, por lo que parece ser un organismo-modelo viable para cumplir con el objetivo de evaluar si las plantas pueden ser organismos sensibles y cognoscentes. Esto porque facilitaría al científico la labor de reconocer en la planta ciertos procesos, tales como respuesta a estímulos, habituación, o aprendizajes por parte de ésta. No obstante, para poder confirmar la existencia de estas capacidades una condición aparentemente necesaria es encontrar mecanismos de transmisión de información similares a los sistemas neuronales (Animal Ethics s.f.): esto es evidenciado en Calvo et al. (2017), quienes dan cuenta de que todas las plantas vasculares tienen un mecanismo nervioso de transmisión de información basado en canales iónicos de calcio y en potenciales de acción, los que son, en último término, mecanismos generadores de señales eléctricas.
Los datos, información y teorías que se puedan generar a través del organismo-modelo debiesen ser aplicables a otros organismos, sobre todo a aquellos que podrían resultar más complejos en algún sentido (Ankeny y Leonelli 2020, 2). Por lo tanto, la información que se pueda desprender del estudio de Mimosa Púdica debiese ser aplicable a otros organismos que resultan ser más lentos o que inician movimientos no visibles para los humanos. La investigación observacional que se desprenda de Mimosa permitirá al investigador establecer una línea base de actividad y niveles de respuesta del modelo (Barnett, 1963). Según Abramson y Chicas-Mosier (2016, 2) la línea base proporciona pistas sobre métodos de recompensa, como los estímulos aversivos y discriminativos, los que pueden incorporarse a los paradigmas de aprendizaje y que pueden entonces ser utilizados para evaluar la existencia de procesos adquisitivos de información en plantas posiblemente mediados por algún tipo de estado mental desconocido (Gagliano 2017).
Analizar la posibilidad sobre si las plantas poseen capacidades sensibles y cognoscentes es una tarea particularmente difícil, ya que estos organismos parecen estar inactivos la mayor parte del tiempo debido principalmente a que los pequeños cambios de comportamiento son difíciles de ver y, por lo tanto,
complejizando la tarea de relacionar dichos comportamientos con las capacidades sensibles y cognitivas (Abramson y Chica-Mosier 2016, 2): por esto Mimosa Púdica podría ser un buen modelo, ya que facilita el reconocimiento de estos procesos desde la perspectiva del investigador. En este caso, la modelo Mimosa Púdica podrá ser utilizada para comprender a otros organismos vegetales a los que no resulta provechoso estudiar directamente a causa de sus respuestas poco accesibles desde la observación directa. Los estudios que puedan desprenderse de este modelo podrán entonces ser utilizados para proyectar la información obtenida más allá del contexto en el que estos fueron producidos (Ankeny y Leonelli 2020, 6) y ser empleados para una mejor comprensión de aquellas características existentes en las plantas que son reconocidas como aptitudes sensitivas y cognitivas.
En el presente trabajo argumentaremos que Mimosa Púdica es un organismo-modelo apropiado para estudiar comportalmente la sensibilidad y cognición en plantas debido a sus rápidos movimientos y cambios en su comportamiento, los que son, a diferencia de la mayoría de las plantas, directamente observables por el observador. Mimosa sería una modelo útil para estos fines pues facilitaría el trabajo del observador en la medición de estos procesos, que se evidenciarían en la respuesta a estímulos, habituación y aprendizaje (Kandel et al. 2014). Para lograr este objetivo, ofreceremos el siguiente recorrido argumentativo
Se realizará un breve recorrido histórico sobre diversas investigaciones en biología, particularmente desde el área de la electrofisiología, las que han utilizado a Mimosa Púdica como organismo-modelo para estudiar la respuesta a estímulos de las plantas
Se argumentará que la respuesta a estímulos es una capacidad propia de organismos sensibles, y que, si bien no todas las plantas inician movimientos visibles, todas son capaces de responder a estímulos debido a mecanismos de transmisión nerviosa (Bose 1926; Calvo et al. 2017).
Y, por último, se postulará que los estudios con Mimosa Púdica han permitido evidenciar ciertos procesos relativos al aprendizaje y habituación a partir de su cambio comportamental y memoria celular (Abramson y Chicas-Mosier 2016), los que son, en último término, características propias de agentes cognitivos (Cisek 2019)
Esto nos permitirá concluir que estudiar estos procesos en nuestro organismo-modelo, tanto en un sentido mecanicista como funcionalista, resulta provechoso para evaluar la presencia de respuestas sensibles y estados cognitivos en plantas: la planta sensitiva permitiría comprender el funcionamiento sensible y cognitivo de las demás plantas
Sensibilidad y estudios de respuesta a estímulos con Mimosa.
Podemos definir sensibilidad como la capacidad de sentir (Calvo et al. 2017, 2859). La cual podemos entender como la capacidad de responder a estímulos. En este sentido, diremos que “responder a estímulos” y “sentir” son equivalentes. La postura de que las plantas son organismos sensibles debe ser defendida a través de estudios electrofisiológicos del análisis sobre las funciones que cumplen los potenciales de acción en organismos vegetales, y también a través de estudios del comportamiento. Respecto a la necesidad de analizar las funciones que tienen los potenciales de acción, esto se debe a que los potenciales de acción (i.e las señales eléctricas) son las encargadas de inducir señales de respuesta ante eventuales amenazas en plantas, y en algunos casos iniciar movimientos que sean visualmente detectables por el ojo humano (Calvo et al. 2017, 2858).
En animales, los potenciales de acción son transportados por las neuronas, las cuales emiten reportes al cerebro sobre el estado de los tejidos del cuerpo e informan sobre la detección de sustancias o situaciones peligrosas o potencialmente peligrosas, las que generan la sensación de dolor (Calvo et al. 2017, 2859). En el caso de las plantas esto puede resultar complejo, pues si bien sabemos que son capaces de emitir señales eléctricas o potenciales de acción (Calvo y Friston 2017), estas no son transmitidas a través de neuronas. No obstante, aquellas señales que en animal causan la sensación de dolor, en plantas inducen
potenciales de acción (Calvo et al. 2017), lo que permitiría al organismo activar algún tipo de mecanismo de defensa como los propuestos por Zavala (2010).
Bose y Guja (1922) descubrieron a través de distintos experimentos con plantas que los potenciales de acción son transportados a través del floema, uno de los dos tejidos vasculares del cuerpo vegetal. El floema, a diferencia del xilema (el otro tejido vascular), es capaz de transportar nutrientes en doble dirección, es decir, tanto desde arriba hacia abajo, como de abajo hacia arriba (Nabors 2006). No es de sorprenderse que estas señales sean transmitidas a través del floema si se toma en consideración que los potenciales de acción en plantas no finalizan su recorrido en un cerebro, sino que más bien se distribuyen a lo largo del sistema. Y eso no es todo, pues el floema, a diferencia del xilema, es un tejido con altos niveles de entrecruzamiento con otros tejidos no-vasculares, lo que es conocido como anastomosis. Esto explica, hasta cierto nivel, como es que las plantas serían capaces de responder a estímulos y generar respuestas, sin importar el lugar desde el que se percibe el estímulo y se inicia la señal (Calvo et al. 2017).
Ejemplos de respuesta a estímulos son presentados a través de distintas investigaciones. Alguno de ellos se presenta en la investigación de Puttonen et al. (2016), en donde se puede apreciar que las hojas de una Betula Pendula cesan sus movimientos al llegar la noche; también se ha visto que las plantas son capaces de responder a estímulos auditivos o vibraciones, aunque ellas no posean pabellón auricular. Es posible que esto se deba a algún tejido que actúe como receptor de vibraciones, como los tricomas (Gagliano et al. 2012). Existen otros casos más llamativos, los que pueden ser observados de manera directa, como el caso de Dionaea muscipula, una planta insectívora que posee una hoja trampa que caza insectos cuando estos se posan en sus hojas (Bohm et al. 2016) o el de Mimosa Púdica, la que cierra sus hojas rápidamente cuando ésta ha sido tocada (Hagihara et al. 2022).
Bose (1926) realizó experimentaciones electrofisiológicas con distintas especies de individuos vegetales, las que consistían en propinar descargas eléctricas a los sujetos de prueba. El autor descubrió que las plantas eran capaces de transmitir dicha descarga eléctrica a lo largo del sistema, y que además la planta generaba su propia señal eléctrica. Evidenció también que todas las plantas que participaron de la experimentación generaron señales eléctricas. Bose propone finalmente que las plantas cuentan con un mecanismo nervioso que permite responder a estímulos, el que funciona con transmisión de impulsos eléctricos. Esto es para Bose un tipo de sistema nervioso vegetal. En diferentes estudios (e.g. Calvo et al. 2017; Calvo y Friston 2017; Shepherd 2012; Volkov 2012; Zimmermann et al. 2016) se concluye que las señales eléctricas son las responsables de inducir respuestas a estímulos en plantas, tal como ocurre con los potenciales de acción en animales. No obstante, Bose fue ampliamente criticado por distintos científicos, especialmente por Burdon-Sanderson, quien afirmaba que solo las plantas capaces de iniciar movimientos visualmente perceptibles por el ser humano producían señales eléctricas (Shepherd 2012). La evidencia científica de los últimos años le ha dado la razón a Bose respecto a la cuestión sobre la generación de señales eléctricas en todo tipo de plantas sin importar si sus respuestas resultan visualmente perceptibles por un observador. Con relación a lo anterior, resulta sugerente postular que todas las plantas son capaces de responder a estímulos a través de este sistema nervioso.
Las críticas al estilo Burdon-Sanderson siguen presentándose en la actualidad por más que aparezcan diversas investigaciones novedosas dispuestas a argumentar a favor de la existencia de capacidades sensibles en plantas. En el blog de animal ethics (s.f) se argumenta que las plantas no se mueven y que por eso serían organismos no sensibles. En vistas a que 1) Mimosa Púdica presenta una respuesta a estímulos iniciada en un periodo corto de tiempo; 2) que puede ser diretamente observada en la disposición de sus hojas (Hagihara et al. 2022); y 3) que ya se ha defendido que las señales eléctricas son las encargadas de las respuestas del organismo; creemos que Mimosa Púdica resulta ser un organismo-modelo práctico para estudiar la sensibilidad en plantas y también un modelo útil para justificar la existencia de mecanismos sensibles en organismos vegetales. Debido a esto, los resultados obtenidos de ella pueden ser proyectables a otros tipos de organismos vegetales, lo que debiese convertir a Mimosa en un modelo apropiado (Ankeny y Leonelly 2020) para los fines que hemos establecido.
Es posible que el primer estudio sobre sensibilidad en plantas empleando una Mimosa Púdica como modelo sea el de Pfeffer (1873), quien evidenció que la estimulación mecánica repetida, tal como tocar repetidamente las hojas, conduce a una disminución de la sensibilidad y, por lo tanto, a una disminución en la cantidad de respuestas. Bose (1906) confirma el descubrimiento de Pfeffer, y aporta a su investigación añadiendo que no tan solo la estimulación mecánica disminuye la sensibilidad de Mimosa, también lo hace la estimulación eléctrica. Las descargas eléctricas propiciadas por Bose a las hojas de Mimosa iniciaban el cierre de sus hojas, y las descargas repetidas imposibilitaban una posterior respuesta. Con esto demostró que un periodo de descanso suficiente era necesario para provocar el cierre de las hojas y que, por lo tanto, el sistema bioeléctrico del organismo juega un rol vital en la señalización y respuestas (Bose 1926; Calvo et al. 2017)
Lo anterior sugiere la siguiente pregunta: ¿Es posible que Mimosa distinga entre tipos de estímulos? La respuesta a esta pregunta podemos encontrarla en Holmes y Gruenberg (1965), quienes realizan un experimento com Mimosa incluyendo una gota de agua y tactos con un dedo como estímulos. En dicho experimento, luego de que la planta se acostumbrara a las gotas de agua, ésta dejo de cerrarse. Una vez habituada, las hojas fueron tocadas con un dedo, y sus hojas se cerraron aun cuando no lo hicieron con las gotas de agua. Este experimento resulta relevante por dos razones (Abramson y Chicas-Mosier 2016, 2): en primer lugar, muestra que la habituación al cierre de las hojas no es producto de una fatiga electrofisiológica; si este fuese el caso, entonces mimosa no hubiese respondido al tacto del dedo. En segundo lugar, este estudio muestra que Mimosa es capaz de discriminar entre estímulos. Lo anterior llevó a Applewithe (1972) a estudiar si las variables que afectan la habituación en animales también afectan la habituación de Mimosa. Descubrió que, si se variaba el intervalo de estímulos, entonces aumentaba el tiempo necesario para la habituación (Abramson y Chicas-Mosier 2016)
De hecho, se ha visto que estímulos sucesivos de impulsos eléctricos en Mimosa reducen la transmisibilidad de los potenciales de acción como producto de la estimulación repetitiva (Calvo et al. 2017, 2863). En animales, esta reducción en la transmisibilidad es conocida como habituación (Kandel et al. 2014). Uno de los estudios más recientes sobre la habituación de Mimosa puede revisarse en Gagliano et al. (2014). En la investigación se utiliza el cierre de las hojas como variable dependiente y la caída vertical del prospecto como estimulo. Emplearon una técnica más controlada e investigaron más fenómenos, incluido el recuerdo a corto y largo plazo y el efecto de la intensidad de la luz (Abramson y Chicas-Mosier 2016): esto confirma los resultados de Holmes y Gruenberg (1965). Posiblemente esta habituación presenciada en Mimosa tenga una correlación directa con el cambio de estado del organismo. El estado de reposo de la planta se ve rápidamente alterado debido al gran procesamiento de estímulos, lo que genera un estado de estrés en la planta y provoca lo que reconocemos como habituación a estímulos. El cambio entre estos dos estados puede evidenciarse en la alteración de la transmisión de frecuencias bio-eléctricas (Bose 1926; Calvo et al. 2017).
Otros ejemplos de habituación pueden observarse en las investigaciones de Pfeffer (1906) con Drosera, o en experimentos realizados por Darwin con la planta Passiflora, citado en Applewhite (1975). Con estos métodos es posible estudiar los comportamientos de las plantas y como es que estas utilizan los potenciales bioeléctricos para producir respuestas a diferentes estímulos provocados por el medio (Abramson 2002).
Es posible que la evaluación de señales y estímulos, como también su priorización, sean producto del sistema excitable propiciado por el tejido vascular implicado: el floema. La red vascular de los organismos vegetales es un sistema complejo e interactivo, que tiene el potencial de evaluar a través de retroalimentaciones generadas a nivel fisicoquímico (Baluska 2010). Ya que las respuestas al estrés ambiental son propiciadas por el sistema vascular, todas las plantas vasculares serían entonces capaces de generar respuestas a estímulos, y Mimosa Púdica puede responder a esos estímulos rápidamente, de forma tal que puede ser observada por los investigadores en cuestión. Esto facilita la evaluación sobre posibilidad de que las plantas respondan a determinado estimulo, que se habitúen, se condicionen, aumenten o disminuyan la alteración de la transmisión de señales, entre otras aptitudes y procesos fisiológicos vegetales.
Hemos presentado hasta aquí diversos estudios que permiten evidenciar capacidades sensibles en plantas, en las que algunas de ellas aprenden, recuerdan, y producen diversos estímulos, pero ¿es esto evidencia de capacidades cognitivas en plantas? Intentaremos responder esto en el próximo apartado desde algunos enfoques cognitivistas.
Comportamiento y características cognitivas en plantas a partir de estudios sobre el aprendizaje y memoria.
La perspectiva tradicional de las ciencias cognitivas establece que el comportamiento puede descomponerse taxonómica y conceptualmente en tres partes (e.g Hurley 2001): percepción, cognición y acción. Estas permitirían construir lo que reconocemos como comportamiento (véase figura 1). La percepción utiliza un primer estadio; la cognición, por su parte, permitiría utilizar en un segundo estadio los datos otorgados por la percepción; y por último la acción, que es aquello que podemos observar de un comportamiento y que utiliza los dos estadios anteriores.
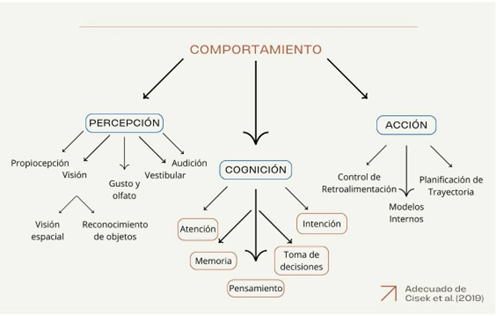
Figura 1. Bosquejo taxonómico conceptual parcial en ciencias cognitivas y neurociencias (Cisek 2019). Tres estadios permiten la aparición del comportamiento: la percepción, la cognición y la acción.
Para esta perspectiva, la percepción es todo aquello que el individuo puede recopilar del medio a través de los sentidos, y la acción es lo que el individuo realiza a partir de los datos ya mediados por la cognición o sus procesos (Cisek 2019, 2)
La cognición es, en un sentido amplio y deflacionario, aquello que permite distinguir entre procesos que utilizan representaciones explícitas del conocimiento y procesos que sencillamente no lo hacen (Cisek 2019, 2). En un sentido más estricto, la cognición se asocia a procesos que generan algún conocimiento a partir de un sustrato material definido: el cerebro (Hommel y Colzato 2015). Este requiere la identificación de cierto sistema a nivel cerebral, el que realiza un conjunto determinado de funciones. Para la perspectiva tradicional, este sistema se encuentra separado del aparato perceptual y de la acción: lo que se interpone entre ambos es lo que se suele llamar “proceso mental” (Hurley 2001; Cisek 2019). Generalmente a la cognición se le asocian diferentes procesos, tales como la toma de decisiones, la intencionalidad, la atención, la memoria y el aprendizaje (Cisek 2019; Abramson y Chicas-Mosier 2016; Kandel et al. 2014). A este propósito, nos centraremos en estudios psicologistas de aprendizaje y memoria en plantas, realizados principalmente en Mimosa Púdica. Estos permitirán argumentar que las plantas pueden ser comprendidas como agentes cognitivas al menos en algún grado.
En Bayne et al. (2019) la cognición se define, en su sentido más amplio, como un acto mental, proceso de conocimiento o habilidad de comprender. El aprendizaje puede ser definido, con relación a la cognición, como una habilidad de comprender que requiere de algún tipo de memoria, al menos de corto plazo: con esta se puede incorporar información externa a un conjunto de información interno (Kandel et al. 2014). Esto es posible en plantas porque, aún cuando no existe un cerebro, existen procesos que permiten la existencia de memoria a nivel celular y que puede ser utilizada por la planta para aprender sobre el medio en el que habita (Abramson y Chicas-Mosier 2016, 3). El aprendizaje es definido por Kandel et al. (2014) como el proceso biológico que permite adquirir nuevo conocimiento sobre el medio en el que habitan los organismos vivos, mientras que la memoria la definen como el proceso de retención de información y reconstrucción del conocimiento.
Se suele asumir que la base de la memoria en sistemas neuronales radica en campos bioeléctricos construidos por células nerviosas (Adey 2004), aunque más recientemente la explicación mecanicista basada en procesos sinápticos se ha vuelto más dominante (Kandel et al. 2014). El énfasis debe realizarse en la noción de “campo”: un sistema integrado, compuesto de movimientos iónicos y cargas eléctricas en la membrana que se construyen a partir de actividades integradas por millones de células que cumplen un papel instructivo en los procesos de crecimiento del organismo y su desarrollo (Calvo et al. 2017, 2864). La construcción del campo implica la existencia de canales iónicos y bombas de membrana que eventualmente modifican el potencial de la membrana plasmática externa, mientras que las proteínas unidas a la membrana suelen asociarse a funciones individuales de la planta (Baluska y Mancuso 2013). Los gradientes bioeléctricos implicados en la transducción de señales son un instructivo epigenético fisiológico que impulsa el crecimiento y la diferenciación (Calvo et al. 2017), y estos están presentes en todas las plantas con sistema vascular (Bose 1926). De hecho, en el artículo de Calvo et al. (2017), se menciona y explica cómo es que las células de los tejidos vegetales pueden almacenar y procesar información si el potencial de la membrana cambia: bajo esta argumentación, podemos decir que las células de los tejidos vegetales actúan como células nerviosas animales y que tienen funcionalmente la misma capacidad.
En la década del 1960, los estudios psicologistas de aprendizaje en plantas se centraron en la posibilidad de aprendizaje sin un sistema nervioso (Holmes y Gruenberg 1965): esto es sumamente importante para los científicos del comportamiento, pues permite determinar si las mismas diferencias en el aprendizaje existentes en vertebrados e invertebrados existen también en plantas (Abramson et al. 2002). Tal como vimos en el apartado anterior, muchos de los estudios tempranos en plantas utilizaron la planta sensitiva como modelo para las investigaciones. De hecho, existen diversas razones para recomendarla como un organismo-modelo óptimo: se sabe sobre su historia natural, son fáciles de mantener y su respuesta (el cierre de las hojas) es fácilmente visible por el investigador.
Sin embargo, el largo tiempo de recuperación puede traer complicaciones, ya que algunas variables de entrenamiento que influyen en el aprendizaje y también el tiempo entre una respuesta y su consecuencia deben ser muy cortas para que pueda establecerse una asociación (Abramson y Chicas-Mosier 2016, 3). Lo anterior implica un segundo problema cuando se comparan las respuestas entre plantas y animales, ya que la velocidad de respuesta es considerablemente más lenta en plantas. Abramson y Chicas-Mosier (2016, 3) mencionan que los potenciales de acción en animales tardan milisegundos en ocurrir, cuando el cierre de las hojas en Mimosa tarda segundos. Debemos diferenciar aquí entre lo entendido como ‘velocidad de la respuesta visible’ (i.e el cierre de las hojas) y la ‘velocidad en que viajan los potenciales de acción’, diferencia que no fue realizada por Abramson y Chicas-Mosier. Un dato a considerar es la velocidad en que viajan las señales eléctricas, y otra es la velocidad en que se genera la respuesta: son dos niveles diferentes y Abramson y Chicas-Mosier los confunden sutilmente. En Volkov (2012) se ha evidenciado que la velocidad de transmisión de los potenciales de acción en plantas no presenta diferencias considerables respecto a la velocidad de transmisión de los potenciales de acción en animales. En este sentido, creemos que la respuesta visible es solo un factor por considerar y que sin duda presenta utilidades al momento de analizar los fenómenos, pero dicho fenómeno no debiese reducirse solo a la respuesta visible. No debemos confundir la velocidad de transmisión de un potencial de acción con la velocidad de generación de respuestas.
En estudios sobre aprendizaje no-asociativo, unos de los más interesantes pueden ser los casos de habituación y sensibilización. La habituación se refiere a una disminución en la respuesta a un estímulo que se presenta repetidamente. La sensibilización, por otro lado, puede considerarse lo opuesto a la habituación, ya que se refiere al aumento en la frecuencia o probabilidad de respuesta (Abramson y Chicas-Mosier 2016, 4). La habituación y la sensibilización resultan adecuadas para el estudio de aprendizaje de plantas: ya que la habituación y la sensibilización se encuentran ampliamente estudiadas en el reino animal, estas proporcionan un marco óptimo para comparar y contrastar los resultados obtenidos en el aprendizaje de las plantas. Lo más importante se desprende de acá, y es que la habituación y la sensibilización comparten muchas propiedades con el aprendizaje que podríamos llamar “más complejo”, como el aprendizaje asociativo o condicionado (e.g Gagliano et al. 2016): esto genera oportunidades únicas para estudiar el comportamiento en plantas. Algunas capacidades, tales como cambios en los patrones de comportamiento, mejora del rendimiento y la sensibilización a partir de estimulación sucesivas (intensidad, frecuencia, capacidad de la respuesta para recuperarse con el tiempo, etc.) puede investigarse fácilmente en organismos vegetales, y la organismo-modelo Mimosa Púdica es, según los estudios presentados en el apartado anterior, una buena alternativa para estudiar estas capacidades.
No obstante, los datos disponibles a la fecha sobre la planta sensitiva no contienen estudios que analicen un amplio espectro de intervalos entre sesiones de entrenamiento. No es posible discriminar entre habituación y adaptación sensorial hasta que dichos datos se encuentren disponibles y correlacionados con los datos previos (Abramson y Chicas-Mosier 2016, 5), aunque investigaciones como la de Hagihara et al. (2022) permiten dar cuenta de que Mimosa es capaz de discriminar entre, por ejemplo, una gota de agua, el tacto de un dedo, y la mordida de un saltamontes. Es sugerente realizar una investigación a futuro que permita analizar y estudiar cómo Mimosa discrimina entre dichos estímulos y aprende a diferenciarlos. Dicha diferencia puede evidenciarse al observar el tipo de respuesta que produce la planta en función del tipo de estímulo (la gota de agua, el dedo o el saltamontes, por ejemplo).
Otro tipo de aprendizaje es el asociativo, y puede observarse en paradigmas de condicionamiento clásico. En éste, una señal conocida como EC (Estimulo condicionado) se entrelaza con un estímulo que provoca un reflejo conocido como EI (Estimulo incondicionado). Después de varios emparejamientos EC-EI, la respuesta provocada por el EI ahora es provocada por el EC (Abramson y Chicas-Mosier 2016, 5; Gagliano 2017; Gagliano et al. 2016). El condicionamiento clásico se demostraría, Según Abramson y Chicas-Mosier, si la planta modelo es capaz de discriminar entre estímulos en relación con su número y patrón de respuestas condicionadas: esto es lo que ocurre en la investigación de Gagliano et al. (2016) con la planta de arvejas o de guisantes Pisum Sativum.
Investigaciones recientes en otros organismos vegetales sustentan la idea de que las plantas puedan adquirir, recordar y procesar información. Un ejemplo podemos encontrarlo en Bohm et al. (2016), quienes realizan análisis bioeléctricos en la planta insectívora Venus Atrapamoscas: la planta es capaz de almacenar información bioelectricamente por periodos cortos de tiempo y puede discriminar entre distintas intensidades de la misma señal eléctrica. Otros ejemplos visualmente menos llamativos podemos encontrarlos en Gagliano (2017) y Gagliano et al. (2016), quienes realizan estudios con la planta de arvejas Pisum Sativum y evidencian la existencia de distintos tipos de aprendizajes, en este caso nos referimos al aprendizaje asociativo, el que implica un almacenamiento de memoria a largo plazo y utiliza distintos estímulos como medios para el aprendizaje. En Gagliano et al. (2016) se utiliza como EC un ventilador, y como EI una luz azul. Numerosos estudios argumentan que la luz azul y la luz violeta son señales deseadas por la planta ya que con ella la planta es capaz de realizar fotosíntesis, la que cumple un rol ecológico determinante para la sobrevivencia del organismo (Trewavas 2009).
Con todo lo anterior mencionado, defendemos que las capacidades cognitivas de las plantas, específicamente el aprendizaje y la memoria, se encuentran ampliamente estudiadas: las plantas serían agentes cognitivos ya que son capaces de aprender y recordar, aunque ello no las convierte en agentes igualmente cognitivas que otros organismos. El problema de afirmar que las plantas son agentes cognitivas reside en las condiciones que se establecen para poder definir un organismo como agente
cognitivo, como lo es con el caso de los animales y la identificación de neuronas. Los organismos vegetales no cuentan con neuronas, pero si con equivalentes funcionales, los que permiten cumplir las mismas funciones. Creemos que los mencionados estudios avalan la afirmación previa
Mimosa Púdica ¿Un buen modelo?
La pregunta que encabeza este apartado ha permitido discutir sobre el estatus epistemológico de varios tipos de modelos producidos por la intervención científica, sea teórica o práctica (Frigg y Hartmann 2018). Lo particular de determinado modelo radica (o debería radicar) en su capacidad representacional de circunstancias epistémicas. En este sentido, un modelo actúa como mediador entre la teoría, la práctica y el mundo (Ankeny y Leonelli 2020, 51). Un modelo puede ser defendido como un buen modelo en tanto herramienta epistémica que permite el acceso a un fenómeno especifico, lo que suele llamarse como “objetivo” o “sistema objetivo” (Target system). Dicha variable epistémica (i.e el acceso al conocimiento), específica del modelo en cuestión, permitiría la proyección de los resultados experimentales a los demás objetos con los que el modelo comparte propiedades.
La problemática radica en otra pregunta: ¿qué hace que tales proyecciones sean más (o menos) plausibles? Esta pregunta no es irrelevante, pues los modelos, y más específicamente los organismos- modelo, generalmente se usan para representar fenómenos que posiblemente no son directamente observables utilizando los propios organismos (Ankeny y Leonelli 2020, 52). Este es el caso de nuestro análisis, pues queremos evidenciar la existencia de sensibilidad y cognición en plantas, y esto no es posible a través de la observación directa en la mayoría de las plantas (el “propio organismo”). Como se adelantó brevemente en el primer apartado, una de las razones por las que se afirma que una planta no puede ser sensible y/o cognoscente, se debe a la información que podemos obtener visualmente de ellas. En este sentido, tenderemos a decir que las plantas no son sensibles ni cognoscentes debido a su nula capacidad de respuesta: concluimos esto debido a la aparente inexistencia de movimiento en ellas. Sin embargo, los mecanismos involucrados en las respuestas, como también en la señalización y mecanismos de defensa, se encuentran ampliamente estudiados y existen en todas las plantas vasculares, independientemente de sus respuestas visibles. De hecho, la evaluación de algunos filósofos de la ciencia sobre lo que constituye un “buen” modelo, se basa en el reconocimiento y articulación de los mecanismos implicados en el fenómeno de interés (Ankeny y Leonelli 2020, 55). Si consideramos que todas las plantas vasculares poseen mecanismos confiablemente similares para la respuesta a estímulos, entonces es posible postular que todas las plantas inician movimientos, pero la velocidad de ellos varía según individuo vegetal. Mimosa serviría para analizar los límites de dichas respuestas y los alcances de las señalizaciones a nivel celular. Si los organismos objetivo (i.e las plantas) comparten características mecanicistas con el modelo, entonces las probabilidades de replicar el fenómeno aumentan, pues las vías causales y sus respuestas ocurrirán de manera similar aún cuando estas vías se alteren levemente. Ankeny y Leonelli (2020, 56) argumentan que los mecanismos subyacentes compartidos por los organismos son la base para establecer lo que podríamos llamar ‘un buen modelo’. La identificación de mecanismos comunes involucrados en el fenómeno de interés (e.g Lee y Calvo 2023) es una parte importante en la evaluación del modelo como un buen o mal modelo.
Algunos han argumentado que las inferencias que se obtienen de un organismo-modelo son extrapolaciones empíricas (Levy y Currie 2015): dichas extrapolaciones implican tratar al modelo como representativo de una clase más amplia de organismos (Bolker 2009), en este caso, apuntamos a las plantas vasculares a nivel general, pero buscamos un punto de facilitación en la evaluación de respuestas a estímulos. Para lo anterior, una planta de respuestas rápidas y visibles es útil, como Mimosa. Es posible proyectar y extrapolar los estudios de sensibilidad y cognición en mimosa al resto de organismos vegetales con sistema vascular debido a mecanismos comunes.
Para finalizar, debemos mencionar que uno de los grandes problemas de la práctica científica con organismos-modelo se debe a la poca explicitación por parte de los científicos del por qué y cuando los organismos-modelo se consideran útiles y apropiados para su uso en la práctica científica (Ankeny y Leonelli 2020, 56). Aquí hemos defendido que Mimosa es una planta modelo óptima para el estudio de
fenómenos sensibles y cognitivos, pues permite evaluar de forma visual y directa sus respuestas a estímulos, aunque esto no implica que no existan ciertas complicaciones en su utilización, tal como señalamos en el apartado anterior. Para comprender a Mimosa como un organismo-modelo apropiado para el estudio de la sensibilidad y cognición en plantas es requisito comprender los procesos comunes implicados en los fenómenos y en el objetivo. En este sentido, se vuelve necesario analizar las características que permiten esta respuesta y los procesos implicados en el fenómeno que se encuentran presentes en todas las plantas vasculares (e.g Abramson et al. 2002; Abramson y Chicas-Mosier 2016; Bohm et al. 2016; Bose 1926; Calvo et al. 2017; Gagliano 2017; Gagliano et al. 2016; Zimmermann et al. 2016), por lo que, en base a la evidencia aquí presentada, todas las plantas son organismos sensibles y agentes cognoscentes en algún grado. La ventaja de Mimosa es que facilita la observación del fenómeno, sin tener que utilizar técnicas de aceleración de fotogramas como los timelapses.